
样本:
二、立论依据
(包括项目的研究意义、国内外研究现状分析,并附主要参考文献及出处)
对基础研究,着重结合国际科学发展趋势,论述项目的科学意义;
对应用基础研究,着重结合学科前沿、围绕国民经济和社会发展中的重要科技问题,论述其应用情景。
虽然人胚胎神经干细胞的研究已经做了大量的工作,然而人们一直在探索成人大脑中是否存在着神经干细胞以及具有那些功能。成年灵长类动物包括人类大脑皮层中是否存在着神经再生现象?是否可从中分离出成年神经干细胞?在2002年正成为神经科学家和干细胞科学家极为关注的问题1、2。人的大脑与认知等高级功能是密切相关的,因此探索在成年大脑中神经细胞的生成对于在生理状态下了解多种神经活动的产生机制和病理状态下脑功能的修复有着极为重要的意义3。
成年神经干细胞的分离和成年神经再生现象有着极为密切的关系4。已知在灵长类动物和人类的海马和脑室下区(SVZ)存在着神经再生现象5、6,是两个干细胞聚集区,2001年从中分离出成人神经干细胞7。然而在大脑皮层却存在着争论1、2、8。Gould于1999年发现猴的大脑皮层存在着神经元再生现象9,这些神经细胞来源于SVZ区,迁徙到大脑皮层并形成突触,9周后凋亡10。Rakic2001年12月的工作提出了反对意见,认为灵长类动物皮层虽然有细胞增生但并不存在神经元再生5。在啮齿类动物研究已证实脑室下区(SVZ)存在着干细胞池,这个区域的干细胞不断地延着嘴侧延长区(rostral extension,RE)向嗅球迁徙,构成了嘴侧迁移流(rostral migratory stream,RMS)11。2000年的研究发现在灵长类也存在RMS5、12。在遵守医学伦理的前提下,我们在2001年从三例开放性脑外伤破碎的脑组织中分离出神经祖细胞13,其中一例破碎脑组织来源于前额叶脑室腹前侧,这一区域类似于灵长类的RE区,分离克隆出的细胞更具有神经干细胞特征。因此,我们推测可能在人类大脑中存在着区别于海马和及脑室下区新的干细胞聚集区,这一部位可能就位于前脑脑室腹侧区14。有趣的是2002年意大利Gritti等15也发现小鼠SVZ的嘴侧延长区(RE)存在着一个新的成年神经干细胞的细胞池,在这个区域中的细胞具有多种干细胞的特性。如果我们的研究工作能够提供充分的实验数据证明我们的设想将是对成人神经干细胞研究的一个重要推动。
同样重要的问题的是成年后新产生的神经干细胞是否具有功能、能否形成突触联系。2002年2月Gage领导的小组首先证实了在小鼠成年新产生的神经元具有确定的功能作用,它可以产生膜电位、动作电位并且能够和周围的细胞形成突触联系3。我们正在详细研究外源性的神经干细胞移植到特定的区域,如海马或侧脑室腹前区是否也能产生功能和突触联系,这对于损伤后干细胞的移植是否具有修复作用及探索其修复机制有着重要的科学意义。
还有一个有待探索的问题是成人神经干细胞和胚胎神经干细胞在增殖和分化潜能上是否相同,如果不同,是否可通过将其细胞核再编程增加发育分化潜能。2002年3月爱丁堡和牛津大学的研究人员在《Nature》上报告将小鼠神经干细胞和胚胎干细胞(ES)混合培养可以产生两种细胞的杂交体细胞,并且具有更加多样的分化潜能16。这一研究有两个重要的启示,首先是干细胞杂交体现象使干细胞的研究中得出有关跨胚层和跨系分化的结论更应谨慎,其可能是两种不同细胞融合的结果17。另一方面通过细胞融合也为干细胞的再编程提供了新的实验手段,虽然其获得的是四倍体杂交细胞,在应用时需谨慎小心,但使神经干细胞具有被改造重塑产生多种潜能的可能。现在还不知道将人的神经干细胞和ES细胞混合培养,是否能融合产生杂交体细胞、以及杂交体细胞发育分化潜能变化和移植到体内有那些生物作用。
二、立论依据
(包括项目的研究意义、国内外研究现状分析,并附主要参考文献及出处)
在将成人神经干细胞应用于临床时应谨慎,要注意免疫排斥问题。我们在进行了大量的体外细胞研究和体内移植实验后,包括在猴脑进行各种毒性安全实验后。经医学伦理委员会同意,在2001年进行了成人神经干细胞自体移植治疗脑损伤的研究,经过术后8个月的随访,观察到一些神经功能的恢复,但这是干细胞移植的作用还是自然恢复的结果需要客观地评估研究。目前已观察到正电子发射断层扫描(PET)显示移植区代谢有所恢复。神经干细胞的研究和应用为人脑再生医学开辟了新的前景。但真正要取得令人信服的证据,表明其再生的功能作用还需进行大量细致的基础神经科学研究。这些研究的进展将推动人类对自身大脑神经生物学认识的飞跃和临床神经科学的发展。
参考文献
1. Rakic P. Neurogenesis in adult primate neocortex: an evaluation of the evidence. Nature Rev Neuroscei, 2002,3:65-71
2. Gould E, Gross CG. Neurogenesis in adult mammals: some progress and problems. J Neurosci ,2002; 22: 619-23
3. van Praag H, Schinder AF, Christie BR, Toni N, Palmer TD, Gage FH. Functional neurogenesis in the adult hippocampus. Nature, 2002, 415: 1030-1033
4. Kintner C. Neurogenesis in embryos and in adult neural stem cell. J Neurosci, 2002, 22: 639-643
5. Kornack DR, Rakic P. Cell proliferation without neurogenesis in adult primate neocortex. Science, 2001, 294: 2127-2129
6. Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C, Peterson DA, Gage FH. Neurogenesis in the adult human hippocampus. Nature Med, 1998, 4: 1313-1317
7. Palmer TD, Schwartz PH, Taupin P, Kaspar B, Stein SA, Gage, FH. Progenitor cells from human brain after death. Nature, 2001, 411: 42-43
8. Arsenijevic Y, Villemure JG, Brunet JF, Bloch JJ, Deglon N, Kostic C, Zurn A,Aebischer P. Isolation of multipotent neural precursors residing in the cortex of the adult human brain. Exp Neurol, 2001, 170: 48-62
9. Gould E, Reeves AJ, Graziano MSA,Gross CG. Neurogenesis in the neocortex of adult primates. Science, 1999, 286: 548
10. Gould E, Vail N, Wagers M, Gross CG. Adult-generated hippocampal and neocortical neurons in macaques have a transient existence. Proc Natl Acad Sci, 2001, 98: 10910-10917
11. Gage FH. Mammalian neural stem cells. Science, 2000 , 287:1433-8
12. Bernier PJ, Vinet J, Cossette M, Parent A. Characterization of the subventricular zone of the adult human brain: evidence for the involvement of Bcl-2. Neurosci Res, 2000, 37: 67-78
13.
14.
15. Gritti A, Bonfanti L, Doetsch F, Gaille I, Alvarez-Buylla A, Lim DA, Galli R, Verdugo JMG, Herrera DG, Vescovi AL. Multipotent neural stem cells reside into the rostral extension and olfactory bulb of adult rodents. J Neurosci, 2000, 22: 437-448
16. Ying QL, Nichols J, Evans EP, Smith AG. Changing potency by spontaneous fusion. Nature, 13 March 2002 (DOI)10.1038/729
17. Terada N, Hamazaki T, Oka M, Hoki M , Mastalerz DM, Nakano Y, Meyer EM, Morel L, Petersen BE, Scott EW. Nature ; 13 March 2002 (DOI )10.1038/730
三、研究方案
1. 研究目标、研究内容和拟解决的关键问题
研究目标:
1、通过荧光流式细胞分离术(FACS)、克隆纯化和细胞动力学实验,摸索成人神经干细胞的最适分离条件,比较分析成人神经干细胞和胚胎神经干细胞的分化潜能和神经递质表型。
2、用逆转录病毒转导绿色荧光蛋白(GFP)标记进行成年神经干细胞起源的空间定位分析并进行神经干细胞形成突触联系的结构和电生理功能研究
3、采用细胞融合技术进行成人神经干细胞再编程研究,观察其表型和分化潜能的变化。
4、进行成人神经干细胞的临床应用研究,探讨干细胞自体移植在脑损伤治疗中的价值。
研究内容
一、成人神经干细胞的分离、克隆和细胞动力学研究
1、采用我们已经建立的三种成人神经干细胞和两株胚胎神经干细胞进行人细胞表面抗原CD1331、花生凝集素(peanut agglutinin, PNA)和热稳定抗原(heat-stable antigen, HSA)的流式细胞分离2,以及用能和DNA结合的荧光染料Hoechst 33342进行干细胞分离。
2、 进行成人和胚胎神经干细胞的克隆分析并进行连续传代的细胞群体动力学研究和多潜能分化研究。
3、进行成人神经干细胞分化后的神经递质表型分析。
二、成年神经干细胞的起源、定位和突触功能研究
1、在弥猴用逆转录病毒转导绿色荧光蛋白(GFP)标记进行进行成年神经干细胞起源的空间定位研究
2、用GFP标记神经干细胞体内移植后切片进行神经细胞电生理的检测和分析
3、用GFP结合辣根过氧化物酶(HRP)标记进行神经干细胞形成突触联系的超微结构观察。
三、神经干细胞细胞核再编程及其表型变化的研究
1、将神经干细胞与胚胎干细胞(ES)的融合形成杂交体细胞
2、融合后进行染色体核型分析并采用RT-PCR进行细胞核再编程基因表达研究,检测融合细胞的UTF13(胚胎干细胞转录协同激活因子)和OCT3/4(POU-domain 转录因子)的表达水平。
3、进行体外细胞和体内移植研究分析融合细胞的增殖和分化潜能。
四、成人神经干细胞的临床应用
1、核磁共振导向的立体定向手术进行成人神经干细胞自体移植治疗脑损伤的研究
2、移植治疗脑损伤后进行正电子发射断层扫描(PET)研究分析干细胞移植治疗效果
3、采用诱发电位分析和脑脊液中检测神经营养因子观察干细胞移植治疗效果。
参考文献:
1. Uchida N, Buck DW, He D, Reitsma MJ, Masek M, Phan TV, Tsukamoto AS, Gage FH, Weissman IL. Direct isolation of human central nervous system stem cells. Proc Natl Acad Sci U S A 2000 ,97:14720-5 .
2. Rietze RL, Valcanis H, Brooker GF, Thomas T, Voss AK, Bartlett PF. Purification of a pluripotent neural stem cell from the adult mouse brain. Nature 2001, 412: 736-9
3. Nishimoto M, Fukushima A, Okuda A, Muramatsu M. The gene for the embryonic stem cell coactivator UTF1 carries a regulatory element which selectively interacts with a complex composed of Oct-3/4 and Sox-2. Mol Cell Biol 1999, 19: 5453-65
2. 拟采取的研究方法、技术路线、实验方案及可行性分析
研究方法
1、流式细胞仪(FACS)分析
细胞经磷酸盐缓冲液(PBS)洗涤后取细胞悬液(4(105 /(l),洗涤细胞两次后加入150ng的PE交联的CD133和FITZ交联的PNA(1:200; Vecta)和HAS(1:200, PharMingen)染色细胞,然后在4(C条件下染色30分钟,洗涤三遍后重新悬浮细胞,加入碘化丙锭(propidium iodide,PI)(100(gml-1; Molecular Probes) 再洗涤三遍后,然后用荧光流式细胞仪(FACScaliburTM,BDIS,San Jose CA)进行分析。分析检测使用Cell Quest软件(BDIS)。选用CD133作为阳性细胞表面标志(CD133+)和PNAlo/HSAlo作为阴性标记来纯化分离细胞。同时我们也联合使用能与细胞DNA结合的荧光染料Hoechst 33342进行干细胞分离。
2、单细胞克隆和长期培养细胞群体分析
球体形成率仅代表了神经干细胞短期培养中的增殖能力,要分析克隆后的神经干细胞长期增殖潜能和其性状的稳定性,因此必需进行细胞群体动力学的检测。将神经干细胞分散成单细胞悬液,在96孔平板中进行克隆培养。将其产生的神经球体再分成单细胞悬液,在60mm平板中培养。每个神经球体分散成单细胞悬液后再产生的球体数目可进行分析比较。同时将球体分成单细胞悬液种植在25cm的培养瓶中(1×105细胞/瓶)。7天传代一次,共5次。每次取一个样本进行检查活细胞数目。分析连续传代1-5次(短期)和15-20次(中长期)神经干细胞群体动力学。
3、逆转录病毒介导的GFP基因转移
因为逆转录病毒只能转染正在分裂的细胞,因此可作为载体将标志物如GFP特异性转导入增殖的细胞中。我们已将GFP标记基因克隆进逆转录病毒载体,我们所使用的是高效泛生型逆病毒载体,逆病毒的包膜基因被水疱性口炎病毒(VSV)编码G蛋白的基因取代,可以通过简单的超速离心办法增加载体的浓度。逆转录病毒转染干细胞在六孔板上进行,载体与靶细胞的比例为1:100,细胞和病毒载体温浴12小时后更换新培养液,继续培养72小时后检测其基因表达的情况。逆转录病毒用于猴脑内干细胞的标记时滴度为1×108 cfu/ml。
4、动物实验
(1)恒河猴,饲养在中科院上海分院灵长类实验动物房(有灵长类动物饲养许可证)。
手术前猴禁食不禁水12小时,用盐酸氯胺酮0.4ml和阿脱品0.5ml肌肉注射后进行麻醉前诱导,待猴进入麻醉状态后用戊巴比妥钠(2.5mg/kg)进行腹腔内注射,待猴进入深麻醉状态后固定于SN-2立体定向头架( NARISHIGE(r) Scientific Instrument Lab)。切开头皮,颅骨钻孔后用Hamilton微量注射器对猴单侧脑室或脑实质注射GFP逆转录病毒(1×107 cpu)或神经干细胞悬液(6 × 104 /(l)到(立体定向坐标如下:A:5.5 mm, R:2.0 mm, H:-2.5 mm, 原点取双侧耳道连线中点上10cm),于2分钟内缓慢进针,停留5分钟后,于3分钟内缓慢退出。然后确切止血后并在伤口撒外用消炎粉,缝合头皮,注射环孢霉素A 10mg/kg。复苏后送回动物房小心饲养,术后8小时禁食。
(2) 新生裸鼠脑室内移植:BALB/c裸鼠,饲养在SPF动物房。新生小鼠在冰中冷冻麻醉2分钟,纤维光源透射显示两侧侧脑室,使用微玻璃纤维注射管将2ul神经干细胞悬液(6×104 cell/(l)注入侧脑室内,在电热毯上复温苏醒后动物放入母鼠笼中,接受抚养,(如将人细胞注入CD-1小鼠,动物每天注射一次环孢霉素A 10 mg/kg,裸鼠则不需要。)细胞移植后2、14、28、56天分别麻醉后处死动物,取脑组织切片进行免疫组化染色。
5、双标记免疫组化
玻片用10%多聚甲醛在室温下固定15mm,用磷酸缓冲液(PBS)洗3次,用blocking液(5% FCS,3% 二抗同源血清,0.3% tritonX-100)室温下固定1小时,然后洗四遍每次15分钟,加入两种一抗,然后再加入FITZ交联的抗鼠IgG二抗37(C温育2小时,然后洗四遍,再用7-amino-4-methylcoumarin-3-acetic acid(AMCA)标记的抗兔IgG二抗体,洗四遍后盖玻前加入荧光盖玻液。
6、超微结构分析
在荧光显微镜下将0.4%的荧光偶连的辣根过氧化物酶(HRP, Sigma)注射到100(m厚切片的每一个GFP+神经干细胞中1,切片经过HRP-DAB处理,在1%的锇酸中固定,然后在环氧化树脂中包埋2。在10000倍的电子显微镜下进行35nm厚连续切面上进行突触分析。
1. van Praag H, Schinder AF, Christie BR, Toni N, Palmer TD, Gage FH. Functional neurogenesis in the adult hippocampus. Nature, 2002, 415: 1030-1033
2. Coleman LA, Friedlander MJ, Intracellular injections of permanent tracers in the fixed slice: a comparison of HRP and biocytin. J Neurosci Methods, 44,167-177(1992)
7、电生理研究
动物大脑移至冷的溶液中(浓度为mM):110胆碱Cl-, 2.5KCL,1.3NaH2PO4, 25.0NaHCO3, 0.5CaCl2, 7MgCl2, 20葡萄糖,1.3Na+抗坏血酸,0.6Na+丙酮酸,5.5犬尿喹啉酸。切片按van Pragg的方法进行,切成约200-400(m厚,然后转移至人造脑脊液中(mM: 125.0 NaCl, 2.5 KCl, 1.3NaH2PO4, 25.0NaHCO3, 2CaCl2, 1.3MgCl2, 1.3Na+抗坏血酸,0.6Na+丙酮酸,10葡萄糖),通以95%O2/CO2(pH7.4, 320mOsm),然后在30?C下。检测脑组织中的GFP+细胞。微电极(6-8MΩ)由硼化玻璃拉制,并充以(mM:120.0 葡萄糖酸钾, 20KCl, 5NaCl, 4MgCl2, 0.1EGTA, 10.0HEPES, 4.0 Tris-ATP, 0.3 Tris-GTP, 10磷酸肌酸,pH7.4, 295mOsm)。细胞膜电位记录采用Axopatch200B(Axon Instruments)并用Labivew-based记录分析软件(L.Campbell, Salk Institute)分析。
8、细胞融合实验和再编程分析
分别将成人和胚胎神经干细胞转导Neo-GFP基因,并将胚胎干细胞转导Hyg基因(Hygromycin磷酸转移酶),将神经干细胞与胚胎干细胞以10:1的比例混合培养,先将细胞置于神经干细胞培养液(DMEM/F12/N2)中两天,然后置于ES细胞培养(EBM-2)条件中,并加入LIF(白血病抑制因子)。将G418(500(g ml-1)加入培养液中4天,再加入Hygromycin(1(g ml-1) 后培养10天,扩增后的细胞用于细胞免疫组化分析、染色体核型分析和RT-PCR。鉴定融合后的细胞中的染色体数目倍数(双倍或四倍)。检测融合细胞的UTF1(胚胎干细胞转录协同激活因子)和OCT3/4(POU-domain 转录因子)表达水平,确定其细胞核再编程的能力和增殖分化潜能。
9、临床神经干细胞移植治疗脑损伤研究
为将神经干细胞精确地移植到脑内特定手术目标点,采用核磁共振扫描导向的立体定向手术进行神经干细胞移植。为避免细胞培养液对细胞移植的影响,神经干细胞被用患者自身的脑脊液洗涤离心后,并悬浮在脑脊液中。立体定向后进行多靶点注射。术前和术后3天、1、6、12月进行CT、MRI(GE, 1.5T)和PET(Siemns ECAT-HR+, 32环,空间分辨率4.2mm)检查。同时检查病人脑脊液中BDNF(ELISA)和EGF(ELISA)以及IL-2、IL-6。每三个月定期进行病人的。从临床中选择年龄、性别、脑损伤程度和部位相似的脑外伤病人进行术后的CT、MRI、PET 、脑脊液和诱发电位检查,进行对照研究。
实验方案
细胞融合实验和再编程实验
1. 采用基因工程制备两种逆转录病毒。一个编码Hygromycin(Hyg)抗药基因,另一个编码新霉素(Neo)抗药基因和GFP基因,两者之间由IRES (internal ribosome entry site)序列分隔,从而可在一个CMV启动子作用下同时表达Neo和GFP基因。
2. 将上述载体质粒和pMD-VSVG质粒用磷酸钙沉淀方法转染293T装配细胞。
3. 收集细胞培养上清,使用超速离心法,纯化收集病毒颗粒。
4. 使用了Neo-GFP逆转录病毒载体转导神经干细胞;Hyg逆转录病毒转导胚胎干细胞。
5. 将神经干细胞与胚胎干细胞以10:1的比例混合培养,
6. 先将细胞置于神经干细胞培养液(DMEM/F12/N2)中两天,
7. 然后置于ES细胞培养(EBM-2)条件中,并加入LIF(白血病抑制因子)。
8. 将G418(500(g ml-1)加入培养液中4天,
9. 再加入Hygromycin(1(g ml-1) 后培养10天。
10. 扩增后的细胞用于染色体核型分析,鉴定融合后的细胞中的染色体数目倍数(双倍或四倍)。
11. RT-PCR分析检测融合细胞的UTF1(胚胎干细胞转录协同激活因子)和OCT3/4(POU-domain 转录因子)表达水平,确定其细胞核再编程的能力。
可行性分析:
1、成人神经干细胞研究比胚胎神经干细胞的研究更为困难,2001年我们已经成功分离成人神经干细胞,为深入研究奠定了坚实的基础。
2、用逆转录病毒转导GFP基因使得我们可以借助于荧光显微镜精确定位移植后的神经干细胞,进行细胞电生理分析和超微结构的研究
3、因为在人脑中研究成人神经干细胞起源的空间定位受到条件限制,采用灵长类动物(弥猴)进行精确的解剖,从而可以对神经干细胞的起源进行空间定位。
4、将神经干细胞与胚胎干细胞进行融合是2002年新发展的一种技术,在胚胎干细胞的作用下神经干细胞的核可以进行再编程,从而有可能改变成年神经干细胞的性状,为探索改造干细胞增殖分化的潜能提供了新的可能。
5、已经进行了成人神经干细胞的自体移植治疗脑损伤积累了初步的临床研究经验。
本项目的创新之处
1、从三例开放性脑损伤患者脑组织中分离出成人神经干细胞,其中一例破碎脑组织来源于前额叶脑室腹前侧,国际上未见报导。从而为在人类大脑中探索新的干细胞聚集区提供了线索。
2、目前国际上尚未有成人神经干细胞移植后的电生理功能和突触超微结构的研究,这一研究对于分析干细胞移植后的修复作用及探索其修复机制有重要的科学意义。
3、采用细胞融合技术进行成人神经干细胞核的再编程是一个新颖的科学研究,虽然其获得的是四倍体杂交细胞,在应用时需谨慎小心,但为改造成年神经干细胞提供了新的技术手段。
4、我们已经进行了世界首例成人神经干细胞的自体移植治疗脑损伤,因此采用客观的实验手段来评价其长期疗效对于临床神经科学具有重要的意义。
年度研究计划及预测进展
2003.1-2003.12
1、进行成人神经干细胞的分离和克隆研究
2、进行群体动力学和细胞动力学的研究
3、开始进行成年神经干细胞起源和定位的研究
2004.1-2004.12
1、进行神经干细胞的电生理和突触功能的研究
2、 进行神经干细胞和胚胎干细胞融合的研究
3、 进行成人神经干细胞的临床应用研究
2005.1 – 2005.12
1、融合细胞的增殖分化潜能变化的研究。
2、成人神经干细胞的临床应用研究
预期研究成果
1、通过对成人神经干细胞分离和克隆的研究,探索高效快捷的分离纯化方法,寻找出神经干细胞分化的递质表型规律,为神经干细胞产业化生产和神经元分化提供重要的理论依据。
2、结合成人神经干细胞的分离和弥猴的实验,探寻灵长类动物的前脑脑室腹侧区是否是一个干细胞新的聚集区,为成年神经干细胞的分离提供一个新的来源。通过对神经干细胞移植后的突触形成的超微结构和电生理功能研究,探讨神经干细胞移植后促进神经功能恢复的修复机制。
3、通过细胞融合技术,使神经干细胞核再编程,从而为增加神经干细胞的增殖分化潜能提供了一个新的途径,对生物技术具有重要意义。
4、我们已经进行了世界首例成人神经干细胞的自体移植治疗脑损伤,通过此研究采用客观的实验手段来分析长期疗效,从而有可能得出在没有免疫排斥条件下干细胞的移植治疗脑损伤治疗价值,为临床应用提供重要的研究基础。
四、研究基础
1. 与本项目有关的研究工作积累和已取得的研究工作成绩
一、成功分离克隆出各种类型神经干细胞
1、2000年5月初,我们在遵守医学伦理及得到医院许可的情况下,将胎龄分别为7周和9周的胎儿流产组织,采用我室所特有的4步法(形态分离、生长因子调节、FACS纯化SP细胞和单克隆化),分离出FZ-B7、FZ-B9二株人胚胎神经干细胞。。另外我们还克隆出CD1小鼠的胚胎神经干细胞和海马成年神经干细胞。
2、采用胚胎眼球组织,我们分离克隆出人视网膜干细胞,其不但具有自我更新能力,而且能分化成光感细胞、双极细胞和米勒胶质细胞。
二、2001年3月开始收治了三例开放性脑外伤病人:两例患者被利器或钝器严重击伤头部,颅骨粉碎、脑浆四溢。外科医生在清创中从头发和骨缝中将破碎的脑组织收集并进行培养。经过克隆和扩增,培养出成人神经祖细胞。另一例患者被锐器异物刺入额叶脑底内深达10公分,抽取出异物后,冲洗异物上附带的碎片脑组织,其碎片脑组织来源于前脑脑室腹前侧,这一区域类似于灵长类的RE区,分离克隆出的细胞更具有神经干细胞特征。为了检验神经干细胞的特征,克隆的神经干细胞在体外进行增殖和分化实验,并移植到免疫缺陷的裸鼠脑内观察其迁徙和分化能力。为了保证临床移植的安全性,还在猴子的脑内进行了神经干细胞移植试验。
三、流式细胞分离(FACS)纯化神经干细胞
我们已成功地使用能与细胞DNA结合的荧光染料Hoechst 33342进行FACS神经干细胞分离。
四、逆转录病毒
我们已将多种标记基因克隆进逆转录病毒载体,我们所使用的是高效泛生型逆病毒载体,逆病毒的包膜基因被水疱性口炎病毒(VSV)编码G蛋白的基因取代,这一改变不仅使载体具有广泛的宿主范围,而且可以通过简单的超速离心办法增加载体的浓度。用于神经干细胞研究的表达GFP基因逆转录病毒的浓度可达1×108cfu/ml。
五、们已将人胚神经干细胞FZ-B9及生长因子转导后的FZ-B9GDNF移植到由6-OHDA诱导的单侧性帕金森氏病恒河猴模型的纹状体中,现已观察到由Apomorphie诱导的旋转试验有所改善。
六、神经干细胞和胚胎干细胞融合实验
为了筛选神经干细胞和胚胎干细胞融合实验后的杂交体细胞,我们使用了两种逆转录病毒。一个编码Hygromycin(Hyg)抗药基因,另一个编码新霉素(Neo)抗药基因和GFP基因,两者之间由IRES (internal ribosome entry site)序列分隔,从而可在一个CMV启动子作用下同时表达Neo和GFP基因。Neo-GFP载体转导神经干细胞;Hyg载体转导胚胎干细胞。将神经干细胞与胚胎干细胞以10:1的比例混合培养,先将细胞置于神经干细胞培养液(DMEM/F12/N2)中两天,然后置于ES细胞培养(EBM-2)条件中,并加入LIF(白血病抑制因子)。将G418(500(g ml-1)加入培养液中4天,再加入Hygromycin(1(g ml-1) 后培养10天,扩增后的细胞用于细胞免疫组化分析、染色体核型分析和RT-PCR。
Primers : Oct3/4:5′-primer TTCCTTCTTGGGTATGGAAT;
3′-primer GAGCAATGATCTTGATCTTC
UTF1: GCCAACTCATGGGGCTATTG; CGTGGAAGAACTGAATCTGAGC
Neuron(PTX3): AGGACGGCTCTCTGAAGAA; TTGACCGAGTTGAAGGCGAA
七、成人神经干细胞自体移植治疗脑损伤的临床应用
我们在进行了大量的体外细胞研究和体内移植实验后,包括在猴脑进行各种毒性安全实验后。经医学伦理委员会同意,在2001年进行了成人神经干细胞自体移植治疗脑损伤的研究,经过术后8个月的随访,观察到一些神经功能的恢复,目前正电子发射断层扫描(PET)显示移植区代谢有所恢复。
2. 已具备的实验条件,尚缺少的实验条件和拟解决的途径(包括利用国家重点实验室和部门开放实验室的计划与落实情况)
一、本实验目前已具备的仪器条件:
(1)、细胞培养常用仪器:CO2培养箱、超净台等, -80(C冰箱、液氮贮存器。
(2)、冰冻切片机、多种抗体和激光双通道共聚焦显微扫描仪和电生理记录仪。
(3)、MAC磁珠分离系统和流式细胞分析仪。
(4)、分子生物学仪器:PCR仪、电泳仪、DNA合成仪、高速离心机、超速离心机。
(5)、SPF动物房和神经外科立体定向动物实验房。
(6)、猴专用脑立体定向仪(NARISHIGE(r) Scientific Instrument Lab.)SN-2型。
(7)、诱发电位仪(Dantec Co. Keypoint IV)
(8)、核磁共振、正电子发射断层扫描(PET,Siemns ECAT-HR+, 32环,空间分辨率4.2mm)
(9)、透射电子显微镜
二、 尚缺少实验条件:需增加生物安全柜Hfsafe-1200(Heal Force Dev. Ltd.)一台
对申请者负责的前一个已结题科学基金资助项目(项目名称及批准号)完成情况,后继研究进展及与本申请项目的关系加以详细说明。另附该已结题项目《研究工作总结摘要》(300字)及已发表主要相关论文首页复印件(限三篇)。
八、申请者承诺
我保证上述填报内容的真实性。如果获得资助,我与本项目组成员将严格遵守国家自然科学基金委员会的有关规定,切实保证研究工作时间,按计划认真开展研究工作,按时报送有关材料。
九、推荐意见
(不具有高级专业技术职务的申请者,须有两名具有高级专业技术职务的同行专家推荐。推荐时,请认真负责地介绍申请者及其项目组成员的业务基础、研究能力、科研态度及研究条件等。项目组成员不能做推荐者)
推荐者(签章) 专业技术职务 专长
所在单位:
推荐者(签章) 专业技术职务 专长
所在单位:
十、申请者所在单位及合作单位的审查与保证
1. 申请者所在单位学术委员会的审查意见(包括:对项目的意义、特色和创新之处及申请者的研究水平与学风签署具体意见)
主任或副主任(签章) 2002年 3 月 21日
2. 合作单位的审查意见与保证
同意参加合作研究,保证对参加合作研究人员时间及工作条件的支持、督促其按计划完成所承担的任务(及需要说明的其它问题)
3. 申请者所在单位领导的审查意见与保证
已按填报说明对申请人进行了资格审查,对申请书内容进行了审核,同意学术委员会的审查意见,并保证在项目获得资助后做到以下几点:
(1) 保证对研究计划实施所需的人力、物力和工作时间等条件给予支持。
(2) 严格遵守科学基金委员会有关资助项目管理、财务等各项规定。
(3) 督促项目负责人和本单位项目管理部门按科学基金委员会的规定及时报送有关报表和材料。
需要说明的其它问题:
单位负责人 (签章) 单位(公章) 年 月 日
全部文件可下载查看
文件下载:
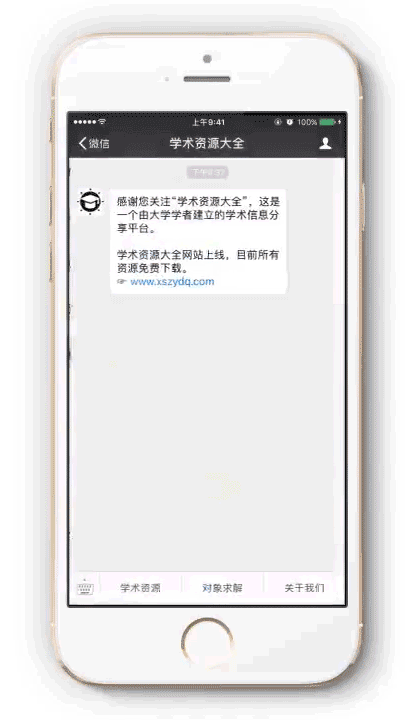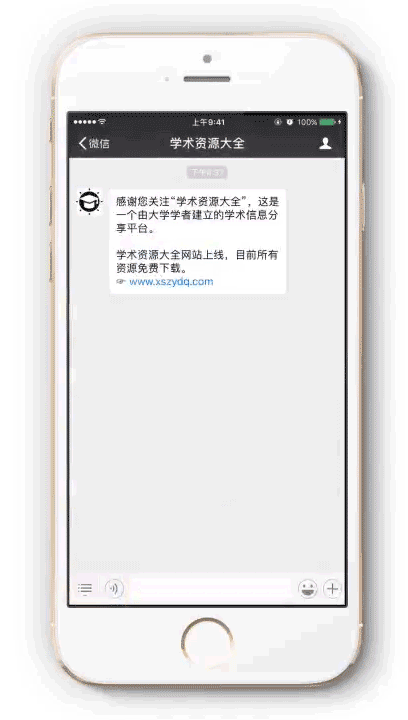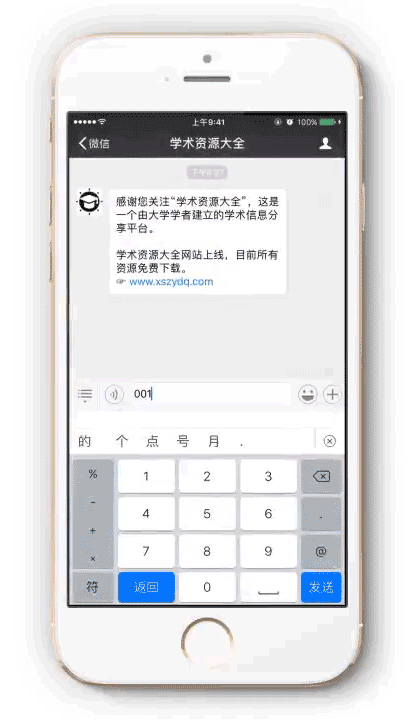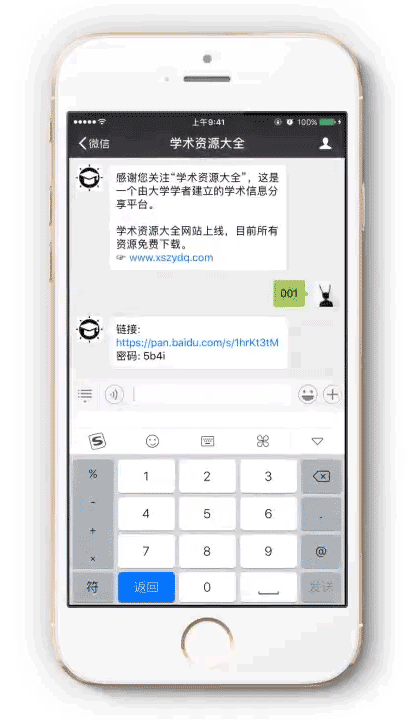
下载地址: http://pan.baidu.com/s/1geK0xNH
提取密码:请按以下步骤获取

1 微信扫描进入公众号
2 向公众号发送文件编号: r0004
3 即可获得提取密码
相关文章
版权声明:
本网站(网站地址)刊载的所有内容,包括文字、图片、音频、视频、软件、程序、以及网页版式设计等均在网上搜集。
访问者可将本网站提供的内容或服务用于个人学习、研究或欣赏,以及其他非商业性或非盈利性用途,但同时应遵守著作权法及其他相关法律的规定,不得侵犯本网站及相关权利人的合法权利。除此以外,将本网站任何内容或服务用于其他用途时,须征得本网站及相关权利人的书面许可,并支付报酬。
本网站内容原作者如不愿意在本网站刊登内容,请及时通知本站,予以删除。